For our international customers, please be advised that orders cannot be placed through our website by customers in countries with International Distributor representation.
Galactose Oxidase - Manual
Source:
Dactylium dendroides
CAS:
9028-79-9
EC:
1.1.3.9
{kind=link}
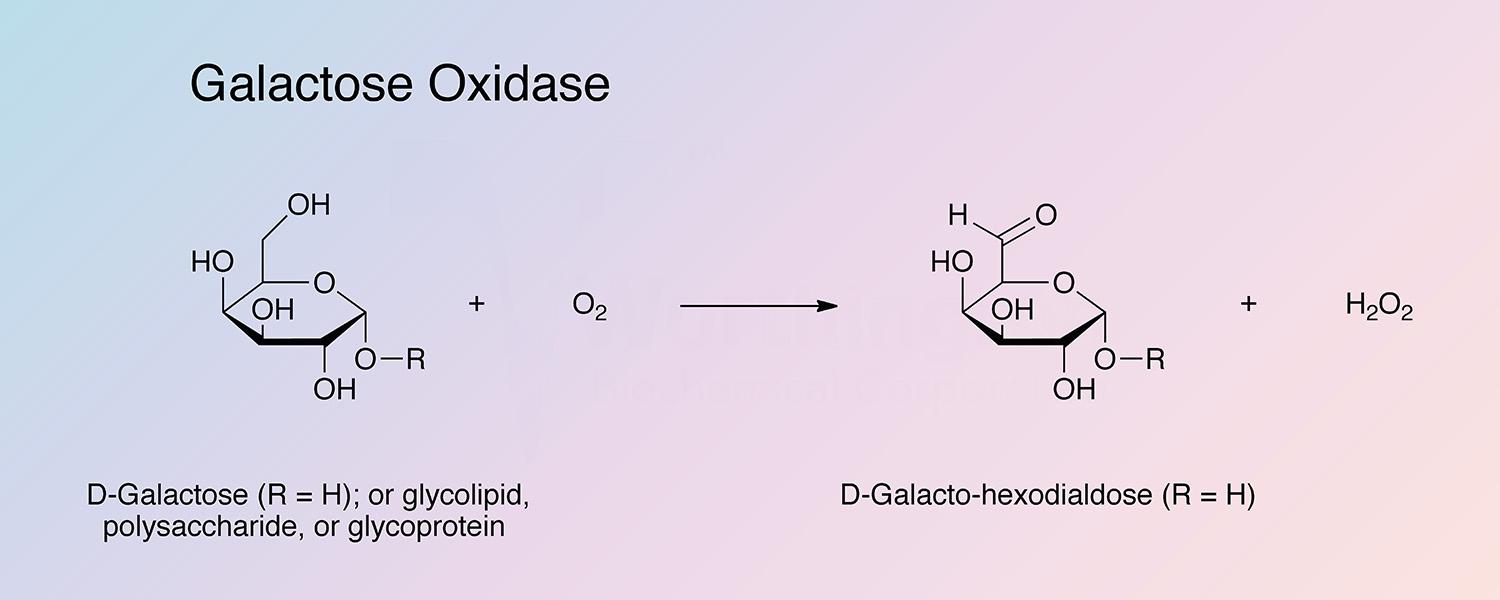
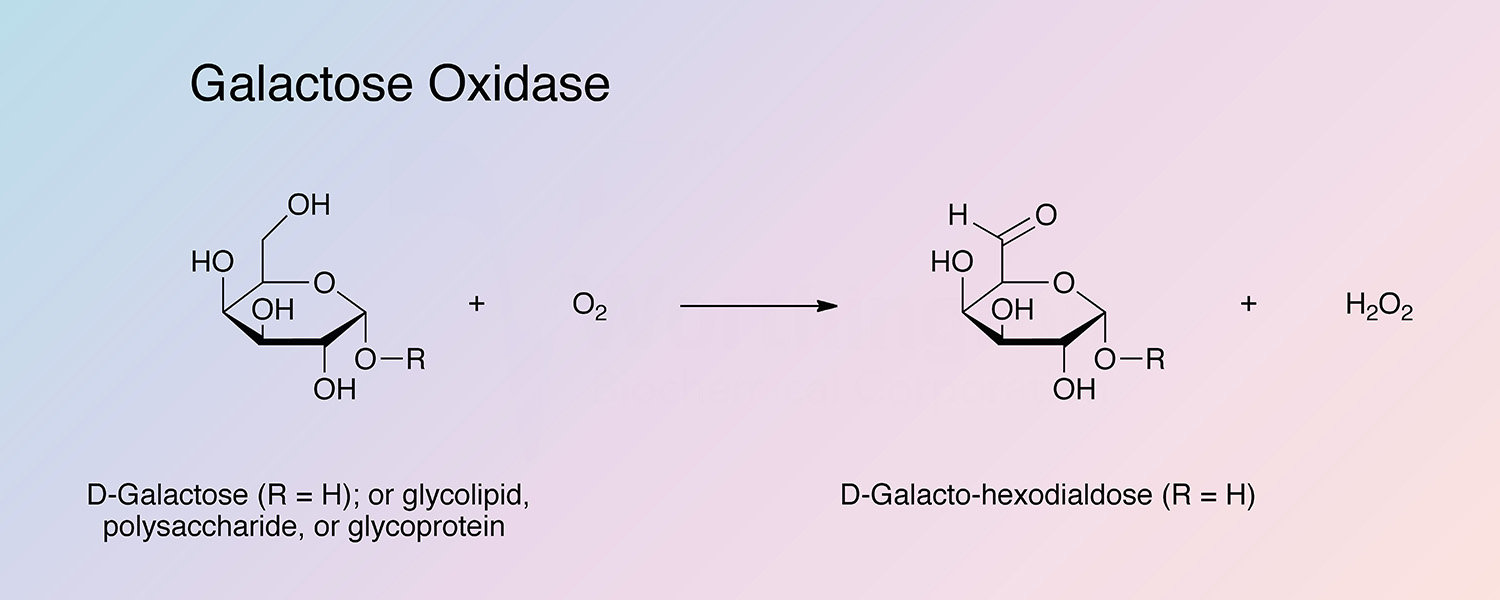
Galactose oxidase (GAO) is a fungal secretory enzyme that catalyzes the oxidation of a range of primary alcohols to the corresponding aldehyde, with reduction of dioxygen to hydrogen peroxide.
History
GAO was discovered in the extracellular culture medium of the fungus Polyporus circinatus by Cooper et al. in 1959. A method to prepare the crystalline enzyme was developed (Avigad et al. 1962, Amaral et al. 1963, Kelly-Falcoz et al. 1965, and Amaral et al. 1966), and it was determined that the original sample was actually Dactylium dendroides and had been misnamed as Polyporus circinatus (Nobles and Madhosingh 1963). Purification procedures were later modified by Bauer et al. in 1967 and Hamilton et al. in 1973.
In the 1980s, the role of carbohydrate content in GAO’s activity was studied (Kornfield and Kornfield 1985, and Medonca and Zancan 1988), and Whittaker did extensive studies on the unique active site of GAO (Whittaker et al. 1988, and Whittaker et al. 1989). Further understanding of the active site was gained through the gene cloning and sequence analysis by McPherson et al. (McPherson et al. 1992, and McPherson et al. 1993).
Recent work with GAO has included successfully cloning and expressing it in E. coli and P. pastoris (Spadiut et al. 2010). An E. coli expression system was also constructed to enable the screening of a library of mutant enzymes (Deacon and McPherson 2011).
Specificity
GAO has a wide substrate specificity, but remarkable stereospecificity, only oxidizing D-isomers of substrates (McPherson et al. 1992). GAO will oxidize galactose and some galactose derivatives in both free and polymeric form. Oxidation occurs at the C6 position.
Composition
GAO contains one Cu(II) atom yet catalyzes a two-electron transfer reaction (McPherson et al. 1992). The copper is bound by two tyrosines, and two histidines (Tyr272, Tyr495, His496, and His581). In a novel post-translational covalent modification, Tyr272 is linked by a thioether bond to cysteine (Cys228), suggesting the involvement of a tyrosine radical in the catalytic mechanism. Stabilization of the radical occurs because Tyr272 of the thioether bond is liganded to the copper, creating a stacking interaction with Trp290 (Whittaker et al. 1989, Ito et al. 1991, and Whittaker et al. 2005). The structure of the enzyme has revealed extensive beta-sheet secondary structure, consistent with the high stability of the enzyme (Kosman et al. 1974).
Most extracellular proteins of eukaryotes are modified by glycosylation during passage through the ER and golgi, leading to greater glycosylation of extracellular than intracellular forms of a protein. Unusually, the intracellular form of GAO is more highly glycosylated (9% carbohydrate) and exhibits greater stability than the extracellular form (2% carbohydrate) (Medonca and Zancan 1988). Additionally, most proteins are modified by O- and/or N-glycosylation while GAO is only modified only by O-glycosylation (Kornfield and Kornfield 1985, and McPherson et al. 1992)
Molecular Characteristics
The gaoA gene contains a long open reading frame from +324 to +2507, including the mature protein-coding sequence (+521 to +2507). It also contains a long untranslated upstream region and a putative pro-sequence with a monobasic cleavage site (McPherson et al. 1992).
Characteristics of Galactose Oxidase
Protein Accession Number
P0CS93
CATH Classification
Three domains:
- Class: Mainly Beta
- Architecture: Sandwich, 7 Propellor
- Topology: Jelly Rolls, Methylamind Dehydrogenase; Chain H, Immunoglobulin-like
Molecular Weight
- 68.5 kDa (calculated from translated DNA sequence and SDS-polyacrylamide gel electrophoresis, McPherson et al. 1992)
- 68.0 ± 3.0 kDa (determined from physical measurements, Cooper et al. 1959)
Optimal pH
- 7.0 (Cooper et al. 1959)
Isoelectric point
- 7.75 (Theoretical)
Extinction Coefficient
- 122,480 cm-1M-1 (Theoretical)
- E1%, 280 = 17.87 (Theoretical)
Inhibitors
- Cyanide
- Diethyldithiocarbamate
- Azide
- Hydroxylamine
- EDTA
- Applications
- Quantitative determination of galactose in blood and other biological fluids (Frings and Pardue 1964, Hankin 1966, and Roth et al. 1965)
- Locating galactose histochemically (Roberts and Gupta 1965)
- Detecting and distinguishing glycoproteins (Itaya et al. 1975)
Galactose Oxidase Products
Description
Activity
Code
Cat. #
Size
Price
Description
Galactose Oxidase
Source:
Dactylium dendroides
Supplied as a lyophilized powder containing sodium phosphate and sucrose as stabilizers.
Store at -20°C. PROTECT FROM MOISTURE.
Ice Pack required
Code
GAO
Product details
LS004520
150 un
$47.00
LS004522
450 un
$82.00
LS004524
1 ku
$140.00
LS004523
Bulk
---