For our international customers, please be advised that orders cannot be placed through our website by customers in countries with International Distributor representation.
Trypsin - Manual
Source:
Bovine Pancreas
CAS:
9002-07-7
EC:
3.4.21.4
{kind=link}
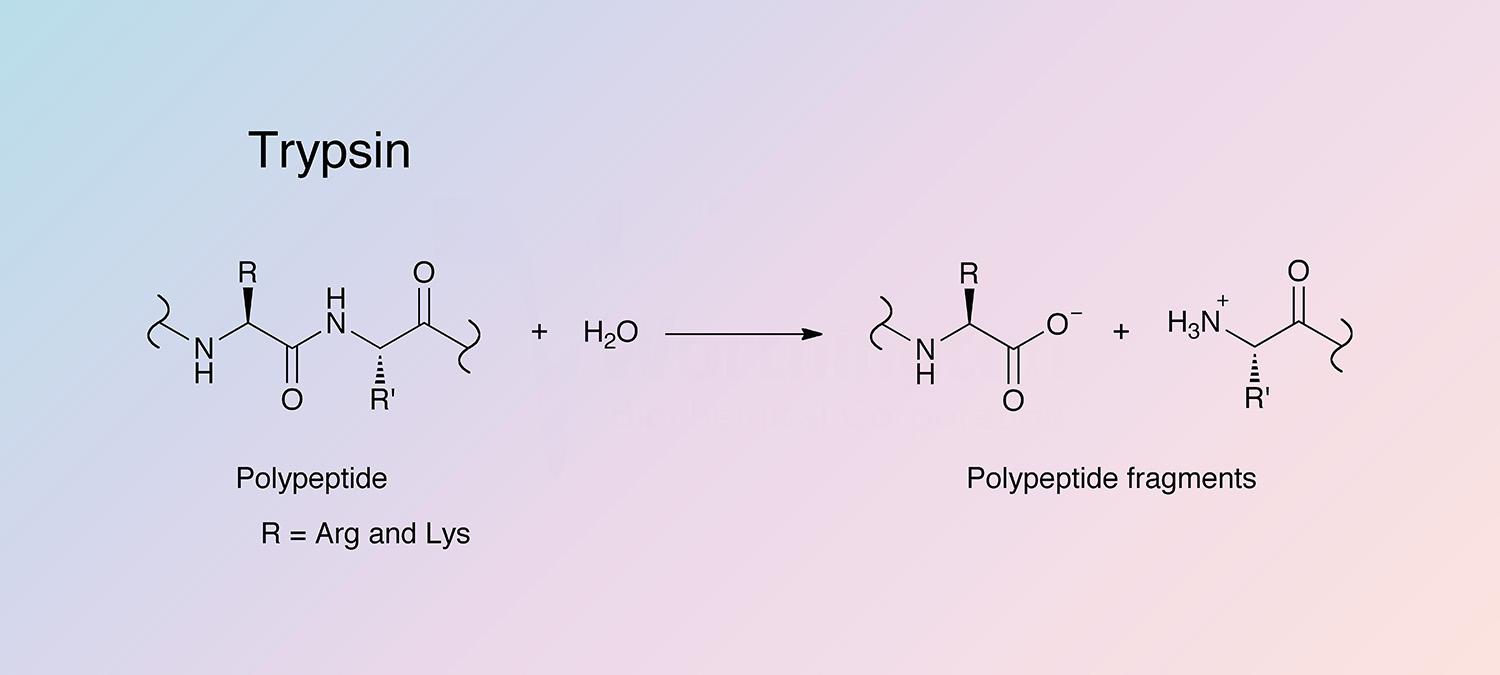
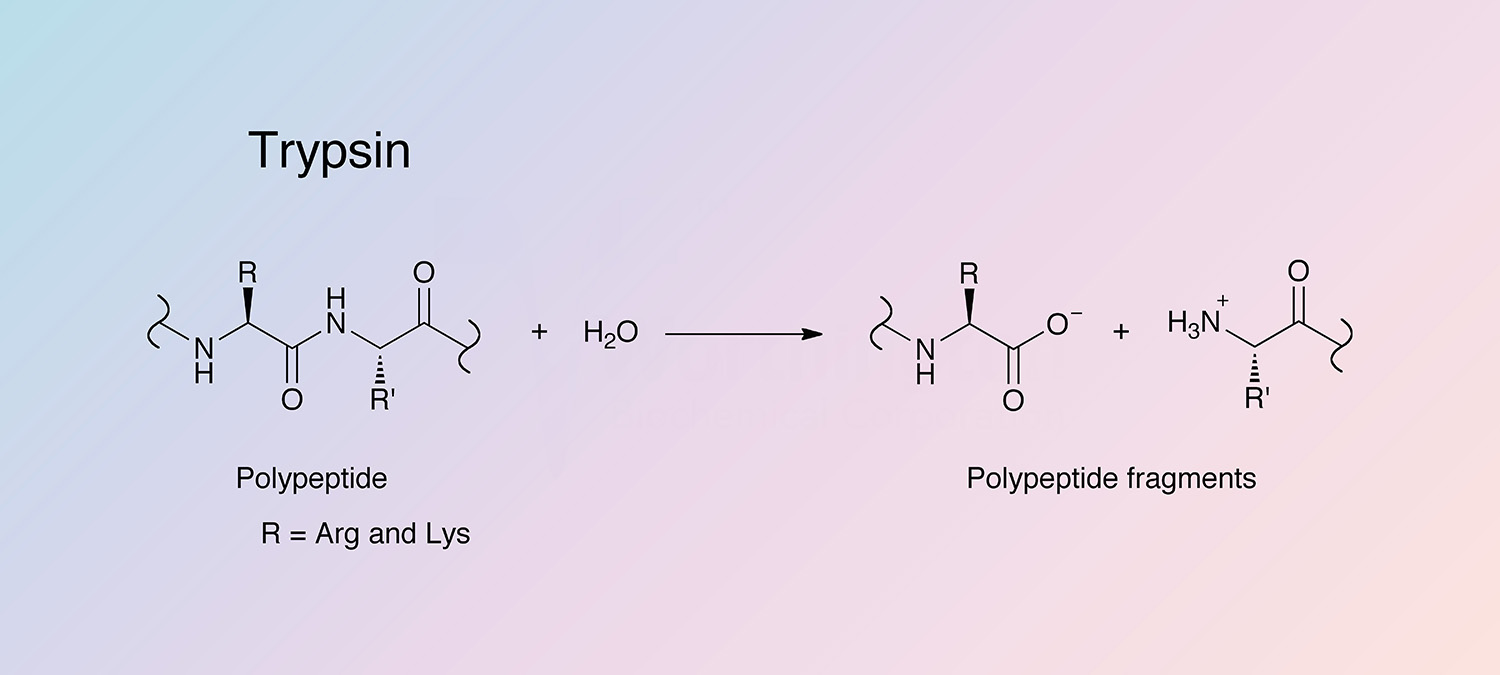
Trypsin is a pancreatic serine protease with substrate specificity based upon positively charged lysine and arginine side chains (Brown and Wold 1973). The enzyme in excreted by the pancreas and takes part in the digestion of food proteins and other biological processes. Trypsin is a medium-sized globular protein and is produced as an inactive proenzyme, trypsinogen (Chen et al. 2009).
History
In 1876, trypsin was first named by Kuhne who described the proteolytic activity of this pancreatic enzyme. He compared trypsin and pepsin, discovering the differentiating factor to be the optimal pH. In 1931, Northrop and Kunitz purified trypsin by crystallization shortly after first purifying pepsin in 1930.
In 1974 the three dimensional structure was determined, which served as a prototype for the serine endopeptidase S1 family to which trypsin belongs.
In the late 1980s, and early 1990s, site directed mutagenesis with recombinant trypsin determined the role of particular amino acid residues (Sprang et al. 1987, McGrath et al. 1989, Corey and Craik 1992, and Corey et al. 1992).
In the late 1990s trypsin’s role in hereditary pancreatitis was investigated, and it was determined that a mutation at Arg117His is responsible for preventing autolysis thereby causing pancreatitis.
Today, trypsin continues to be used in the development of cell and tissue culture protocols (Soleimani et al. 2009, Banumathi et al. 2009, and Yang et al. 2009), as well as protein identification through peptide sequencing techniques (Manz et al. 2004 and Schuchert et al. 2009). In the medical field, the role of trypsin in pancreatic diseases, including cystic fibrosis (Tzetis et al. 2007, and Li et al. 2009) and chronic pancreatitis (Chen et al. 2009), has been the subject of current research, and trypsin has been used to model the decomposition of articular cartilage in osteoarthritis (Wang et al. 2008).
Specificity
Trypsin cleaves peptides on the C-terminal side of lysine and arginine amino acid residues. If a proline residue is on the carboxyl side of the cleavage site, the cleavage will not occur. If an acidic residue is on either side of the cleavage site, the rate of hydrolysis has been shown to be slower.
Composition
Trypsinogen may be activated by removal of a terminal hexapeptide to yield single-chain β-trypsin. Subsequent limited autolysis produces other active forms having two or more peptide chains bound by disulfide bonds. The predominant forms are α-trypsin, having two peptide chains and β-, a single chain. Different activity and thermal stability are shown by α- and β-trypsin.
Other structural features include surface loops at amino acids 185-193, which influence specificity, despite not making direct contact with the substrate. A high affinity Ca2+ binding site is required for stability, and when not present, autolysis occurs. The autolysis loop (located at amino acids 143-151) is very flexible in both trypsin and trypsinogen. Cleavage at the lysine yields the alpha form which retains some catalytic activity. The protein has six completely conserved disulfide bonds (Halfon and Craik 1998).
Molecular Characteristics
Bovine pancreas expresses two forms of trypsin, the dominant cationic and minor anionic forms. These protein sequences share 72% identity, while their coding regions share 78% identity.
Each of these proteins are further processed into alternate forms. Catalytic trypsin contains a flexible “autolysis loop” (residues G145-V157) (Schroeder and Shaw 1968, and Bartunik et al. 1989), and autolysis of the dominant, single-chain form B-trypsin at K148-S149 within this loop leads to the formation of A-trypsin. Further autolysis at K193-D194 leads to the formation of Psi-trypsin (Fehlhammer and Bode 1975).
Both the cationic and anionic trypsin proteins are expressed as trypsinogen proenzymes, with a 15-residue signal peptide (M1-A15) and an 8-residue propeptide (F16-K23). The three-dimensional fold of all known trypsins is highly conserved. In addition, the catalytic triad and regions flanking the catalytic triad are highly conserved (Hartley 1970).
Applications
- Tissue dissociation, especially when combined with other enzymes such as collagenase, and elastase
- Cell harvesting by “trypsinization”
- Mitochondria isolation
- in vitro studies of proteins
- Removing monolayers of cells from plastic and glass
- Various hemagglutination procedures
- Sample preparation for flow cytometric DNA analysis
- Tryptic mapping
- Fingerprinting and sequencing work
- Environmental monitoring
- Reduction of cell density in tissue culture
- Subculturing cells
- Cleavage fusion proteins
- Generating glycopeptides from purified glycoproteins
(White and White 1997)
Characteristics of Trypsin
Protein Accession Number
P00760
CATH Classification
- Class: Mainly Beta
- Architecture: Beta Barrel
- Topology: Thrombin, subunit H
Molecular Weight
23.3 kDa (Theoretical)
Optimal pH
7.5-8.5 (Koutsopoulos et al. 2007)
Isoelectric point
- Trypsinogen: pH 9.3 (Walsh and Neurath 1964)
- Trypsin: pH 10.5 (Cunningham 1954).
Extinction Coefficient
Trypsinogen: 45,250 cm-1 M-1>
Active Site Residues
- Histidine (H63)
- Aspartic acid (D107)
- Serine (S200)
Activators
The rate of trypsinogen conversion is enhanced by using lanthanide in place of calcium ions (Gomez et al. 1974).
Inhibitors
- Pancreatic-, soybean-, lima bean-, and egg white- trypsin inhibitors (see section on Trypsin Inhibitors)
- DFP
- Aprotinin
- Ag+
- Benzamidine
- EDTA
(White and White 1997)
Trypsin Products
Description
Activity
Code
Cat. #
Size
Price
Description
SequENZ® Trypsin, Modified, Sequencing Grade
Source:
Bovine Pancreas
Trypsin, treated with L-(tosylamido-2-phenyl) ethyl chloromethyl ketone to inhibit contaminating chymotryptic activity, chemically modified to promote stability and further purified to remove autolysis fragments, resulting in a highly stable trypsin product resistant to autolysis while retaining specificity.
Store at -20°C. PROTECT FROM MOISTURE.
Ice Pack required
Activity
≥150 units per mg protein (at least 8,625 BAEE/2875 USP/NF units per mg protein)
Code
TRSEQZ
Product details
LS02120
4x25 µg
$98.00
LS02122
4x100 µg
$275.00
LS02123
1 mg
$500.00
LS02124
Bulk
---
Description
SequENZ® Trypsin, Modified, Sequencing Grade, Solution
Source:
Bovine Pancreas
Ready to use liquid preparation of Trypsin, treated with L-(tosylamido-2-phenyl) ethyl chloromethyl ketone to inhibit contaminating chymotryptic activity, chemically modified to promote stability and further purified to remove autolysis fragments, resulting in a highly stable trypsin product resistant to autolysis while retaining specificity. Supplied as 1mgP/ml in 1.0mM HCl.
Store at 2-8°C.
Ice Pack required
Code
TRSEQZS
Product details
LS02150
250 ug
$130.00
LS02152
1000 ug
$415.00
LS02155
Bulk
---
Description
Trypsin, Purified, Sequencing Grade II
Source:
Bovine Pancreas
Bovine trypsin that has been treated with L-(tosylamido-2-phenyl) ethyl chloromethyl ketone (TPCK) to inhibit contaminating chymotryptic activity and extensively purified to remove autolysis products. Supplied as a lyophilized powder.
Store at -20°C. PROTECT FROM MOISTURE.
Ice Pack required
Activity
≥150 units per mg protein (at least 8,625 BAEE/2875 USP/NF units per mg protein)
Code
TRSEQII
Product details
LS02115
4x25 µg
$88.00
LS02117
4x100 µg
$235.00
LS02119
1 mg
$420.00
LS02118
Bulk
---
Description
Trypsin, TPCK Treated
Source:
Bovine Pancreas
A chromatographically purified, diafiltered, lyophilized powder that has been treated with L-(tosylamido-2-phenyl) ethyl chloromethyl ketone (TPCK) to inhibit contaminating chymotryptic activity [Kostka, V., and Carpenter, F.: JBC, 239, 1799 (1964)].
Store at 2-8°C. PROTECT FROM MOISTURE.
Activity
≥180 units per mg protein (10,350 BAEE/3,450 USP/NF units per mg protein)
Code
TRTPCK
Product details
LS003740
100 mg
$118.00
LS003741
500 mg
$420.00
LS003744
1 gm
$705.00
LS003742
Bulk
---
Description
Trypsin
Source:
Bovine Pancreas
Supplied as a chromatographically purified, diafiltered and lyophilized powder.
Store at 2-8°C. PROTECT FROM MOISTURE.
Activity
≥180 units per mg protein (10,350 BAEE/3,450 USP/NF units per mg protein)
Code
TRL3
Product details
LS003708
100 mg
$55.00
LS003707
1 gm
$340.00
LS003709
Bulk
---
Description
Trypsin
Source:
Bovine Pancreas
Supplied as a dialyzed and lyophilized powder.
Store at 2-8°C. PROTECT FROM MOISTURE.
Activity
≥180 units per mg protein (10,350 BAEE/3,450 USP/NF units per mg protein)
Code
TRL
Product details
LS003702
100 mg
$30.00
LS003703
1 gm
$160.00
LS003704
10 gm
$1,150.00
LS003706
Bulk
---
Description
Trypsin, TPCK-Treated, Irradiated
Source:
Bovine Pancreas
Chromatographically purified trypsin treated with L-(tosylamido-2-phenyl) ethyl chloromethyl ketone (TPCK) to inhibit contaminating chymotryptic activity according to Kostka, V., and Carpenter, F.H.: JBC, 239, 1799 (1964), Code: TRTPCK, lyophilized, irradiated and tested for the absence of mycoplasma and extraneous virus according to 9 CFR 113.53c. Each vial is filled to contain 100 mg.
Store at 2-8°C.
Activity
≥180 units per mg protein (10,350 BAEE/3,450 USP/NF units per mg protein)
Code
TRTVMF
Product details
LS003750
100 mg
$180.00
LS003752
5x100 mg
$725.00
Description
Trypsin, 2X, Sterile, Irradiated
Source:
Bovine Pancreas
Purified trypsin (Code: TRL), lyophilized, irradiated and tested for the absence of mycoplasma and extraneous virus according to 9 CFR113.53c. Each vial is filled to contain 100 mg.
Store at 2-8°C.
Activity
≥180 units per mg protein (10,350 BAEE/3,450 USP/NF units per mg protein)
Code
TRLVMF
Product details
LS004454
100 mg
$116.00
LS004452
5x100 mg
$420.00
LS004458
Bulk
---
Description
Trypsin, 0.22µ Filtered
Source:
Bovine Pancreas
Trypsin chromatographically purified, diafiltered (Code TRL3) filtered thorugh a 0.22 micron pore size membrane and lyophilized in sterile vials. This product is not tested for pyrogenicity.
Store at 2-8°C.
Activity
≥180 units per mg protein (10,350 BAEE/3,450 USP/NF units per mg protein)
Code
TRLS
Product details
LS003736
50 mg
$54.00
LS003734
5x50 mg
$235.00
LS003738
Bulk
---